Please use the form below to submit comments. Also provide an e-mail address and name. Your e-mail address and/or name will be used only to communicate with you about this or future comments you may submit. I am particularly keen to receive references to published material that contradicts the assertions and arguments I have made.
By submitting the above comment, I grant to Ross Alan Hangartner the right to incorporate the comment in full or in part, literally, paraphrased, or conceptually, as he sees fit, into State of Pain or other writings that he may create in the future. However, I don't grant permission to include my name or e-mail address, or to use them in any other way than to contact me for follow-up. I understand that by submitting the comment I acquire no right of any kind in State of Pain or other writings of Ross Alan Hangartner.
Last updated: Wed, Jun 21, 2017
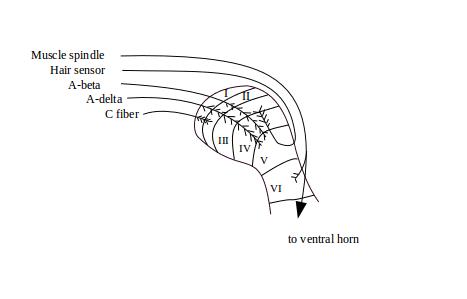
We saw earlier (Three Principal Types of Sensory Neurons) that there are several classes of sensory neurons. They vary in a number of ways including the thickness of their axons. The thicker the sensory fiber, the more deeply it penetrates the dorsal horn. The largest and most rapidly-conducting sensory fibers are those that originate with muscle spindle receptors and tendon strain receptors. These penetrate to lamina VI and continue on into the ventral horn, where they synapse directly with muscle-activating neurons.
A-beta fibers end in laminae III, IV and V. These are large rapidly-conducting fibers that signal stimulation of the various innocuous touch and temperature sensors that densely cover the skin. After these fibers enter the dorsal horn, they split into ascending and descending fibers that emerge from the gray matter and may travel as far as several segments. As they extend, they put out side branches that re-penetrate the dorsal horn, and eventually terminate (arborize) in dense branches of terminals within laminae II through V. This allows the spinal cord to coordinate inputs and outputs across the segments of the spinal cord.
Small lightly myelinated A-delta fibers terminate mainly in laminae I and II, although some extend to lamina V. The A-delta fibers carry signals from both nociceptive and innocuous receptors. A-delta fibers also split into ascending and descending branches. In most cases these branches ascend and descend in a tract along the outside of lamina I called Lissauer's tract. In some cases, the ascending fibers exit the gray matter and ascend in the white matter, putting out side branches that re-penetrate the gray matter. In contrast to the muscle spindle, hair sensor, and A-beta fibers, these arborize through the depth of the dorsal horn, from lamina I through V.
Unmyelinated C fibers, which are largely nociceptive, penetrate only as far as lamina 2. These fibers also split and ascend and descend along Lissauer's tracts, putting out side branches that arborize in laminae I and II.
Figure 1: Arborization of afferents in the dorsal horn1 shows the areas of arborization of the various classes of sensory fibers. This is a cross-sectional view, so of course it does not show the extension of the fibers up and down the spine.
A-delta and C sensory fibers arborize in the same laminae regardless of whether they carry innocuous or nociceptive signals. There is no specialized structure in the spinal cord for processing exclusively painful stimuli.